It is one of two primary mechanisms, along with glycogenolysis, used to maintain a proper glucose level in our blood. It also provides glucose when the dietary intake is insufficient or during stress, such as physical activities or starvation.
Pathway of Gluconeogenesis
Regulation of Gluconeogenesis
Compare and Contrast Gluconeogenesis with Glycolysis and Glycogenolysis
Steps
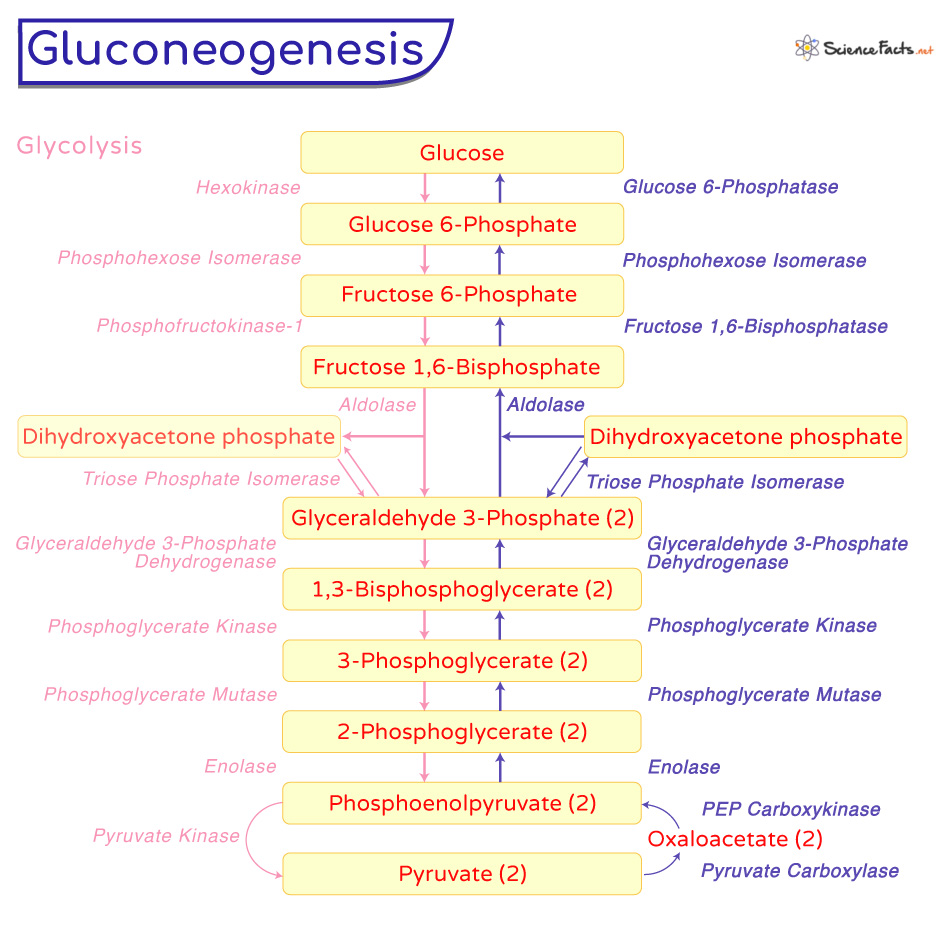
The pathway starts from pyruvate and ends with the formation of glucose. It is a pathway of 11 enzyme-catalyzed reactions. Depending on the substrate used, the path begins in the mitochondria or cytoplasm. Although pyruvate is the primary substrate of gluconeogenesis, all citric acid cycle intermediates (through oxaloacetate), amino acids like alanine, and glycerol also act as a substrate. Steps 1 and 2: Pyruvate to Phosphoenolpyruvate The conversion of pyruvate to phosphoenolpyruvate occurs in two steps, with the formation of oxaloacetate as the intermediate. The first step occurs in the mitochondrion, where pyruvate is carboxylated to form oxaloacetate by the enzyme pyruvate carboxylase. The enzyme requires ATP for energy and biotin as a coenzyme. As oxaloacetate cannot pass through the mitochondrial membranes, it is first converted to malate by malate dehydrogenase. Malate then crosses the mitochondria membrane into the cytoplasm. It again forms oxaloacetate through another malate dehydrogenase. In the second step in the cytosol, oxaloacetate is decarboxylated and rearranged to form phosphoenolpyruvate (PEP) by the enzyme PEP carboxykinase. The enzyme requires GTP as the energy source and Mg2+ as the cofactor. These two steps are unique to gluconeogenesis as they are required to bypass the irreversible reaction catalyzed by the enzyme pyruvate kinase in glycolysis. Step 3: Phosphoenolpyruvate to 2-Phosphoglycerate PEP is hydrated to form 2-phosphoglycerate by the enzyme enolase. Step 4: 2-Phosphoglycerate to 3-Phosphoglycerate In this step, 2-phosphoglycerate is rearranged to 3-phosphoglycerate with the help of the enzyme phosphoglycerate mutase. Step 5: 3-Phosphoglycerate to 1,3-Bisphosphoglycerate In the fifth step, 3-phosphoglycerate is phosphorylated through the enzyme phosphoglycerate kinase to form 1,3-bisphosphoglycerate. This step requires ATP. Step 6: 1,3-Bisphosphoglycerate to Glyceraldehyde 3-Phosphate Here, 1,3-bisphosphoglycerate is reduced to glyceraldehyde 3-phosphate by the enzyme glyceraldehyde 3-phosphate dehydrogenase. In this step, NADH acts as the electron donor. Step 7: Glyceraldehyde 3-phosphate to Dihydroxyacetone phosphate The glyceraldehyde 3-phosphate is then isomerized to dihydroxyacetone phosphate through the enzyme triose phosphate isomerase. Step 8: Glyceraldehyde 3-Phosphate and Dihydroxyacetone Phosphate to Fructose 1,6-Bisphosphate In this step, glyceraldehyde 3-phosphate and dihydroxyacetone phosphate are combined to form fructose 1,6-bisphosphate through the enzyme aldolase. Step 9: Fructose 1,6-Bisphosphate to Fructose 6-Phosphate Fructose 1,6-bisphosphate is further dephosphorylated to form fructose 6-phosphate with the help of the enzyme fructose 1,6-bisphosphatase. It is a rate-limiting step of the pathway. Like the first and the second step, this is also unique to gluconeogenesis, which bypasses the irreversible reaction catalyzed by the enzyme phosphofructokinase-1 in glycolysis. Step 10: Fructose 6-phosphate to Glucose 6-Phosphate Fructose 6-phosphate is further converted to glucose 6-phosphate by phosphohexose isomerase. Step 11: Glucose 6-phosphate to Glucose It is the final step of gluconeogenesis. It involves the conversion of glucose 6-phosphate to glucose through dephosphorylation of glucose 6-phosphate by the enzyme glucose 6-phosphatase. Like the other three steps, this step is also unique to glycolysis, bypassing the irreversible reaction catalyzed by the glycolytic enzyme hexokinase. The factors that regulate the substrates, enzymes, and reactions involved in gluconeogenesis are:
- The conversion of pyruvate to PEP is regulated by acetyl-CoA.Acetyl CoAactivates pyruvate carboxylase through allosteric modification. When concentrations of acetyl-CoA are high, organisms use pyruvate carboxylase to channel pyruvate away from the TCA cycle. It prevents the simultaneous occurrence of gluconeogenesis and the TCA cycle in the cells. Thus, acetyl CoA acts as the indicator of the cells’ metabolic activity.
- The conversion of fructose-1,6-bP to fructose-6-P through fructose-1,6-phosphatase is negatively regulated and inhibited by the molecules AMP and fructose-2,6-bP. AMP and fructose-2,6-bP positively regulate Phosphofructokinase. Again, when there is a high ATP/AMP ratio, the organism increases gluconeogenesis and decreases glycolysis. The opposite happens when during a low ATP/AMP ratio when the organism increases glycolysis and decreases gluconeogenesis.
- The conversion of glucose-6-P to glucose is regulated by substrate-level phosphorylation. As levels of glucose-6-P increase, glucose-6-phosphatase increases activity, and more glucose is produced, thus preventing glycolysis.
- Glucagon regulates gluconeogenesis by reducing the levels of fructose-2,6 bisphosphate, an allosteric regulator.
Phosphorylation of pyruvate kinase causes its inactivation (a covalent modification)Increasing the expression of PEP carboxykinase by glucagon through CRE response elements
5.Availability of substratessuch as pyruvate, glycerol, and amino acids other than lysine (mainly alanine) 6. Adenosine monophosphate (AMP) allosterically inhibits fructose-1,6 bisphosphatase The ratio of ADP/ATP must be meager for gluconeogenesis as it is an energy-dependent pathway. Hormones like glucagon, growth hormone, epinephrine, and cortisol also stimulate gluconeogenesis. In contrast,
Glycolysis is the catabolic breakdown of glucose to form pyruvate that occurs in the cytosol.Glycogenolysis is the breakdown of glycogen in the liver when blood glucose levels drop. It occurs in the liver, muscles, and adipose tissue.