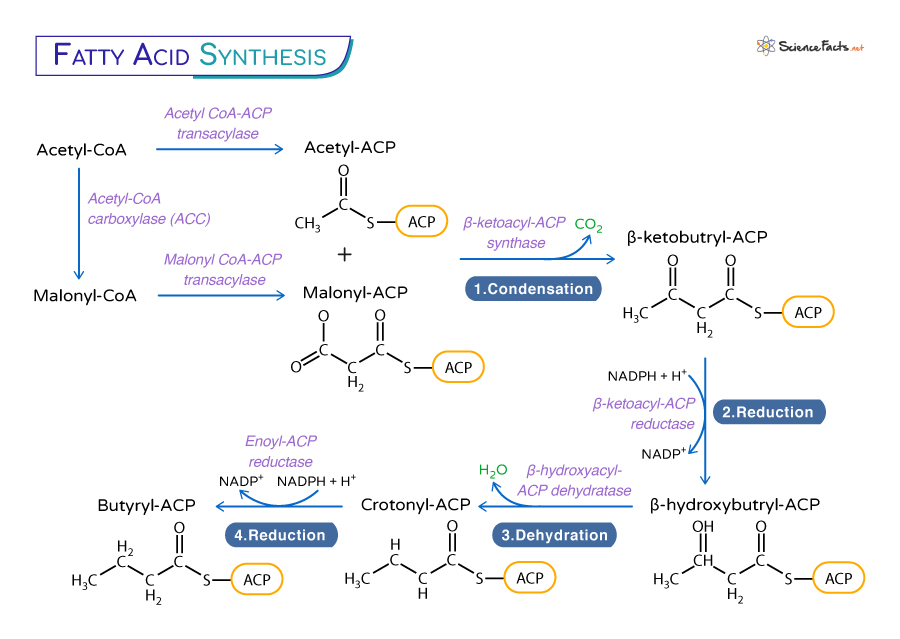
Fatty Acid Synthesis Pathway
Palmitic Acid Synthesis
Synthesis of Long-Chain Fatty Acids
Desaturation of Fatty Acids
Synthesis of Acetyl-CoA
The pyruvate formed through glycolysis is then transported to mitochondria, where it is converted to acetyl-CoA by the enzyme pyruvate dehydrogenase (PDH) complex. The synthesized acetyl-CoA from glycolysis and β-oxidation of fatty acids is transported from the mitochondria to the cytoplasm through a shuttle system. In the first step, acetyl-CoA reacts with oxaloacetate to give citrate, which gets transported from the mitochondria to the cytosol. The citrate is then cleaved back to oxaloacetate and acetyl-CoA with the hydrolysis of one ATP molecule.
Formation of Malonyl-CoA
Once acetyl-CoA is available in the cytoplasm, one molecule is carboxylated to malonyl-CoA by the enzyme acetyl-CoA carboxylase (ACC) with the help of a carbon dioxide molecule and a molecule of ATP. ACC requires biotin as a cofactor. Then, both acetyl-CoA and malonyl-CoA molecules have their CoA replaced by an acyl-carrier protein (ACP), by acetyl CoA-ACP transacylase and malonyl CoA-ACP transacylase, respectively forming acetyl-ACP and malonyl-ACP. ACC is the only regulated enzyme in fatty acid synthesis and thus catalyzes the rate-limiting step of the pathway.
Fatty Acid Chain Elongation
It involves four successive steps involving the repeated addition of two-carbon units to a growing fatty acid chain. Fatty acid synthase (FAS), a large multienzyme complex, carries out the chain elongation steps of fatty acid biosynthesis.
Condensation: It is the first step of fatty acid elongation. Acetyl-CoA and malonyl-CoA are joined together by β-ketoacyl-ACP synthase (also known as the condensing enzymes), resulting in a four-carbon molecule known as β-ketobutryl-ACP with the release of a carbon dioxide molecule. Reduction: Acetoacyl-ACP is then reduced by NADPH to form β-hydroxybutryl-ACP by the enzyme β-ketoacyl-ACP reductase.Dehydration: The β-hydroxybutyryl-ACP further undergoes dehydration, removing a water molecule to create a double-bonded crotonyl-ACP. The enzyme β-hydroxyacyl-ACP dehydratase catalyzes this step. Reduction: The double-bonded fatty acid chain is further reduced by NADPH to yield a saturated acyl-ACP, the butyryl-ACP, by the enzyme enoyl-ACP reductase.
These four steps constitute one round of the fatty acid synthesis cycle. The number of cycles determines the length of the synthesized fatty acid chain. In bacteria, these activities are found on separate enzymes.
Termination
Once the desired fatty acid length is obtained, the growing fatty acid chain is released from the FAS due to the thioesterase activity of the FAS complex. It can then undergo additional modifications, such as saturation of any remaining double bonds or incorporation into triglycerides for storage. The de novo palmitate synthesis requires seven fatty acid elongation cycle rounds. A 16-carbon chain is attached to the ACP arm (palmitoyl ACP) at the end of seven cycles. The 16 palmitoyl ACP is hydrolyzed to release palmitate. The net reaction of palmitate synthesis is given below: Acetyl CoA + 7 Malonyl CoA + 14 NADPH +14 H+ → Palmitate + 7 CO2 + 8 CoA + 14 NADP+ + 6H2O Desaturated fatty acids are rarely found in animals due to their inability to catalyze reactions beyond 9 and 10 carbons. Thus, humans cannot synthesize linoleic acid, which is provided as a dietary supplement.